GENOMA HUMANO
Genoma y genes:
Llamamos genoma al conjunto de todo el ADN de una célula de una especie y los genes que éste contiene. En sentido estricto, el genoma humano no sólo comprende al ADN del núcleo sino también al de las mitocondrias que, aunque sólo tiene 16.000 bases de longitud, es esencial para el funcionamiento celular. Los genes son segmentos de ADN capaces de ser transcriptos –es decir, copiados– a una molécula de ARN (ácido ribonucleico) con igual secuencia que el gen. Los genes no se encuentran yuxtapuestos a lo largo de los cromosomas, sino más bien esparcidos y separados a grandes distancias por secuencias de ADN intergénicas. Las regiones intergénicas constituyen el 70% del genoma, mientras que los genes representan sólo un 30%. Se estima que el genoma humano tiene unos 20.000 genes. Estos genes codifican distintos tipos de ARN, entre los que se encuentran los llamados ARNs mensajeros, que codifican a su vez proteínas. Los otros ARNs, los que no son mensajeros, reciben el nombre genérico de ARNs no codificantes: no son intermediarios entre el gen y la proteína sino que cumplen funciones en sí mismos. Entre éstos están los ARNs ribosomales, de transferencia, nucleares pequeños, los micro ARNs y las ribozimas. Por lo tanto, la definición según la cual un gen es el segmento de ADN que codifica una proteína, no es estrictamente correcta: muchos genes codifican proteínas, pero no todos. Cada uno de los genes que codifican proteínas tiene regiones que estarán representadas en el ARN mensajero maduro intercaladas por otras cuyas secuencias no estarán representadas allí. Las primeras regiones se llaman exones, en tanto que las segundas son los intrones. Mientras los intrones no son codificantes, la mayoría de los exones son las regiones del genoma que codifican proteínas. Estas regiones constituyen sólo el 1,5% del genoma.
Cada cromosoma tiene muchos genes, y la posición que ocupa cada gen a lo largo del cromosoma se denomina locus (del latín, lugar). Cada gen tiene entonces su copia homóloga en el locus equivalente del otro cromosoma del par. Cada una de las dos copias del gen se llama alelo.
Digamos, entonces, que una célula humana tiene dos alelos para cada uno de sus 20.000 genes distintos. No todos los genes se expresan (es decir, se transcriben y se traducen) al mismo tiempo y en el mismo lugar. En un tejido o tipo celular determinado se expresa un subconjunto del conjunto de todos los genes. Uno de los puntos clave de la regulación de la expresión de los genes, es el control de la transcripción. Este control no sólo se ocupa de “encender” o “apagar” genes (efecto del todo o nada), sino también de regular la cantidad de producto (ARN o proteína) de los genes “encendidos”.
Sorprendentemente, el 50% de nuestro genoma está formado por secuencias repetidas en su mayoría de origen viral. Muchas de estas secuencias, mayormente localizadas en regiones intergénicas y en intrones, corresponden a los llamados elementos móviles, o transposones, segmentos de ADN "saltarines", es decir, que pueden duplicarse e insertarse en otras regiones dentro del mismo genoma.
Splicing:
La enzima que transcribe cada gen fabrica un ARN precursor, llamado transcripto primario, que copia tanto los exones como los intrones. Dentro del núcleo un sistema enzimático elimina los intrones del transcripto primario y une sus exones entre sí. Finalmente, el ARN mensajero maduro sin intrones abandona el núcleo y llega al citoplasma donde el proceso de traducción, llevado a cabo por los ribosomas, permite fabricar la proteína correspondiente. Un mismo gen puede dar muchas variantes de proteína. El mecanismo se conoce como splicing alternativo, y consiste en que durante el splicing algún exón, por ejemplo, pueda ser alternativamente incluido o excluido del ARN mensajero maduro. Se estima que podría haber centenares de miles de proteínas distintas codificadas por los 20.000 genes del genoma humano. Esto evidencia que el desciframiento del genoma no es suficiente para entender en su real complejidad el funcionamiento de las células en el plano molecular y que será necesario acelerar la etapa del estudio de las proteínas, en lo que se denomina la era posgenómica o de la proteómica. Por otra parte, como el número de genes humanos no es significativamente mayor que el de un gusano o el de una mosca, se estima que la mayor prevalencia del splicing alternativo en los humanos y otros vertebrados explica su mayor complejidad.
Genotipo y fenotipo:
Al conjunto de la información genética particular de un individuo lo llamamos genotipo. El genotipo es esencialmente la secuencia de ADN. Todo aquello que “vemos” y que no es secuencia de ADN es el fenotipo (del griego, fainein, visible). El fenotipo es tanto lo macroscópico (forma, anatomía, fisiología, patología y comportamiento), como lo más pequeño (histología, bioquímica y estructura molecular). El fenotipo siempre es el resultado de la interacción de un determinado genotipo con un determinado ambiente, lo cual se expresa con la fórmula:
FENOTIPO = GENOTIPO + AMBIENTE
Para algunos fenotipos, la influencia del componente genético es mayoritaria o determinante y, en consecuencia, la ambiental es virtualmente nula. Un buen ejemplo son las enfermedades hereditarias como la fibrosis quística, la corea de Huntington y la distrofia muscular: quien haya heredado el gen de la distrofina mutado padecerá la enfermedad, no importa en qué ambiente se encuentre. En otros fenotipos, la influencia del componente ambiental es mayoritaria y la del genético despreciable. Las enfermedades infecciosas producidas por contagio son un ejemplo.
Para la mayoría de los rasgos fenotípicos hay tanto un componente genético como uno ambiental. Muchas veces estamos seguros de la existencia de ambos pero no conocemos, o incluso sabemos que es difícil de estimar, la contribución parcial de cada uno. La mayor parte de las enfermedades no estrictamente hereditarias, como las cardiovasculares, los cánceres, la hipertensión, el asma, el Alzheimer, el Parkinson y las autoinmunes como la artritis y el lupus, pueden tener un componente hereditario, pero sin duda poseen un componente ambiental no despreciable y, a veces, preponderante. En muchos casos la genética molecular ha llegado a descubrir genes cuyas mutaciones provocan predisposición a dichas enfermedades, pero eso no significa que quien tenga esa mutación padecerá indefectiblemente la enfermedad.
El comportamiento humano también es consecuencia de la interacción del genotipo con el ambiente. Sin embargo, salvo en unos pocos trastornos neurológicos hereditarios, no se ha demostrado una influencia genética específica. La pretendida base genética de fenotipos tan complejos como la inteligencia, la orientación sexual, la criminalidad, las capacidades artísticas o deportivas, debe ser tomada con pinzas y sujeta a riguroso análisis experimental en cada caso particular. De lo contrario se corre el riesgo de caer en el determinismo genético, que lejos de ser una ley biológica es un instrumento de discriminación y dominación socioeconómica.
El conocimiento del genoma no autoriza a nadie a estigmatizar a las personas como resultado irreversible de lo que ordenan sus genes. Los genes nos dicen que podemos hablar, pero no qué idioma; que podemos amar, pero no a quién; que podemos disfrutar de la música, pero no de cuál.
Los distintos tipos de inteligencia, las capacidades, los afectos y nuestros actos son resultados del proceso de culturización, el cual no está registrado en ningún gen y, en cambio, está fuertemente influenciado por el ambiente familiar, social y económico en que vivimos.
Lo heredado y lo adquirido:
Para referirnos al carácter heredado o adquirido de un rasgo resulta muy importante diferenciar tres conceptos biológicos: congénito, genético y heredable. Congénito es aquello que le puede ocurrir al embrión o al feto durante la vida intrauterina, sea o no causado por mutaciones en los genes (es decir, genético) y sean o no esas posibles mutaciones heredadas de alguno de los padres (heredable). Así, un defecto o característica congénita de un individuo puede provenir de situaciones vividas por la madre durante el embarazo o simplemente de fenómenos no controlables conocidos como “ruido” del desarrollo embrionario. En este caso, afectará la vida de ese individuo pero no se transmitirá a su descendencia.
Una característica genética está causada por alteraciones en los genes, pero no es necesariamente heredable. Por ejemplo, un tumor de piel tiene origen genético porque está producido por mutaciones en los genes de alguna célula de la piel. Pero ese cambio en los genes de esa célula no es transmitido a la descendencia porque no afecta a las células germinales (óvulos o espermatozoides). Quiere decir que es genético, pero no es heredable.
Por último, lo heredable, que siempre es genético, es lo único que podría ser tenido en cuenta para avalar una teoría puramente determinista. Las alteraciones genéticas se transmiten mediante ciertas leyes de padres a hijos.
Cuando sin pruebas se atribuyen de comportamiento o capacidades intelectuales humanas a los genes, y se supone que las variantes de estos genes están distribuidas en forma asimétrica en distintos grupos humanos, se terminan postulando hipótesis deterministas como, por ejemplo, que ciertos grupos tienen un techo intelectual y que no “vale la pena” invertir dinero en educación para ellos porque están “genéticamente” limitados.
Genoma, variabilidad genómica y razas:
Podemos definir una especie como un conjunto de individuos capaces de dar descendencia fértil por reproducción sexual. Los individuos que pertenecen a una misma especie a menudo presentan diferencias genéticas. Esto es así porque, aunque para cada gen el individuo presenta dos alelos –dos variantes del mismo gen–, el número de alelos de un gen existente en la especie en su conjunto es generalmente mayor que dos. Un gen puede tener decenas de alelos diferentes, pero en cada individuo sólo habrá dos de ellos. Una raza es una subpoblación de individuos de una especie que tiene una alta homogeneidad genética, es decir que los individuos que la componen comparten muchos más alelos entre sí que con cualquier otro individuo de la misma especie, pero de otra raza.
Las comparaciones de secuencias de ADN entre humanos indican que las grandes diferencias genéticas, de existir, tienen lugar entre individuos y no entre poblaciones. En términos más sencillos, por ejemplo, una persona caucásica (blanca) de Europa puede compartir muchas más variantes alélicas con un asiático o un africano que con otro europeo del mismo color de piel. Dos negros africanos pueden distar mucho más genéticamente entre sí que cualquiera de ellos respecto de un blanco. Esto hace, por ejemplo, que en muchos casos la histocompatibilidad (o sea, cuán compatibles son sus órganos y tejidos) entre un negro y un blanco sea mayor que entre dos individuos de la misma “raza” y, como consecuencia, un negro sea más apto que un blanco para donar un órgano de trasplante a otro blanco. Estudios moleculares del genetista Svante Paabo confirman que las razas humanas no existen. El mismo concepto fue expresado en forma sencilla y elegante por el genetista brasileño Sérgio Pena: “No es que seamos todos iguales, sino que somos todos igualmente distintos”.
Una de las fuentes de variabilidad intraespecífica son los cambios de una base en la misma posición entre distintos individuos. Estos cambios son llamados SNPs (pronunciado snips), por single nucleotide polymorphisms. Si bien los SNPs no están uniformemente distribuidos a lo largo de nuestro genoma, se estima que aparece en promedio un SNP cada mil bases de secuencia. Esto indica que el genoma de nuestra especie, Homo sapiens, es altamente homogéneo con una similitud de secuencia del 99,9%. Las similitud de secuencia entre humanos y chimpancés es de alrededor del 98%. No obstante, hay diferencias, tanto entre humanos entre sí como entre humanos y chimpancés que no se deben a SNPs sino a variaciones del número de copias de un dado segmento de ADN dentro del mismo genoma, lo cual puede elevar las diferencias entre individuos a niveles del 5-6%.
Aún no está claro cómo contribuyen las variaciones en el número de copias al fenotipo de los humanos.
Llamamos genoma al conjunto de todo el ADN de una célula de una especie y los genes que éste contiene. En sentido estricto, el genoma humano no sólo comprende al ADN del núcleo sino también al de las mitocondrias que, aunque sólo tiene 16.000 bases de longitud, es esencial para el funcionamiento celular. Los genes son segmentos de ADN capaces de ser transcriptos –es decir, copiados– a una molécula de ARN (ácido ribonucleico) con igual secuencia que el gen. Los genes no se encuentran yuxtapuestos a lo largo de los cromosomas, sino más bien esparcidos y separados a grandes distancias por secuencias de ADN intergénicas. Las regiones intergénicas constituyen el 70% del genoma, mientras que los genes representan sólo un 30%. Se estima que el genoma humano tiene unos 20.000 genes. Estos genes codifican distintos tipos de ARN, entre los que se encuentran los llamados ARNs mensajeros, que codifican a su vez proteínas. Los otros ARNs, los que no son mensajeros, reciben el nombre genérico de ARNs no codificantes: no son intermediarios entre el gen y la proteína sino que cumplen funciones en sí mismos. Entre éstos están los ARNs ribosomales, de transferencia, nucleares pequeños, los micro ARNs y las ribozimas. Por lo tanto, la definición según la cual un gen es el segmento de ADN que codifica una proteína, no es estrictamente correcta: muchos genes codifican proteínas, pero no todos. Cada uno de los genes que codifican proteínas tiene regiones que estarán representadas en el ARN mensajero maduro intercaladas por otras cuyas secuencias no estarán representadas allí. Las primeras regiones se llaman exones, en tanto que las segundas son los intrones. Mientras los intrones no son codificantes, la mayoría de los exones son las regiones del genoma que codifican proteínas. Estas regiones constituyen sólo el 1,5% del genoma.
Cada cromosoma tiene muchos genes, y la posición que ocupa cada gen a lo largo del cromosoma se denomina locus (del latín, lugar). Cada gen tiene entonces su copia homóloga en el locus equivalente del otro cromosoma del par. Cada una de las dos copias del gen se llama alelo.
Digamos, entonces, que una célula humana tiene dos alelos para cada uno de sus 20.000 genes distintos. No todos los genes se expresan (es decir, se transcriben y se traducen) al mismo tiempo y en el mismo lugar. En un tejido o tipo celular determinado se expresa un subconjunto del conjunto de todos los genes. Uno de los puntos clave de la regulación de la expresión de los genes, es el control de la transcripción. Este control no sólo se ocupa de “encender” o “apagar” genes (efecto del todo o nada), sino también de regular la cantidad de producto (ARN o proteína) de los genes “encendidos”.
Sorprendentemente, el 50% de nuestro genoma está formado por secuencias repetidas en su mayoría de origen viral. Muchas de estas secuencias, mayormente localizadas en regiones intergénicas y en intrones, corresponden a los llamados elementos móviles, o transposones, segmentos de ADN "saltarines", es decir, que pueden duplicarse e insertarse en otras regiones dentro del mismo genoma.
Splicing:
La enzima que transcribe cada gen fabrica un ARN precursor, llamado transcripto primario, que copia tanto los exones como los intrones. Dentro del núcleo un sistema enzimático elimina los intrones del transcripto primario y une sus exones entre sí. Finalmente, el ARN mensajero maduro sin intrones abandona el núcleo y llega al citoplasma donde el proceso de traducción, llevado a cabo por los ribosomas, permite fabricar la proteína correspondiente. Un mismo gen puede dar muchas variantes de proteína. El mecanismo se conoce como splicing alternativo, y consiste en que durante el splicing algún exón, por ejemplo, pueda ser alternativamente incluido o excluido del ARN mensajero maduro. Se estima que podría haber centenares de miles de proteínas distintas codificadas por los 20.000 genes del genoma humano. Esto evidencia que el desciframiento del genoma no es suficiente para entender en su real complejidad el funcionamiento de las células en el plano molecular y que será necesario acelerar la etapa del estudio de las proteínas, en lo que se denomina la era posgenómica o de la proteómica. Por otra parte, como el número de genes humanos no es significativamente mayor que el de un gusano o el de una mosca, se estima que la mayor prevalencia del splicing alternativo en los humanos y otros vertebrados explica su mayor complejidad.
Genotipo y fenotipo:
Al conjunto de la información genética particular de un individuo lo llamamos genotipo. El genotipo es esencialmente la secuencia de ADN. Todo aquello que “vemos” y que no es secuencia de ADN es el fenotipo (del griego, fainein, visible). El fenotipo es tanto lo macroscópico (forma, anatomía, fisiología, patología y comportamiento), como lo más pequeño (histología, bioquímica y estructura molecular). El fenotipo siempre es el resultado de la interacción de un determinado genotipo con un determinado ambiente, lo cual se expresa con la fórmula:
FENOTIPO = GENOTIPO + AMBIENTE
Para algunos fenotipos, la influencia del componente genético es mayoritaria o determinante y, en consecuencia, la ambiental es virtualmente nula. Un buen ejemplo son las enfermedades hereditarias como la fibrosis quística, la corea de Huntington y la distrofia muscular: quien haya heredado el gen de la distrofina mutado padecerá la enfermedad, no importa en qué ambiente se encuentre. En otros fenotipos, la influencia del componente ambiental es mayoritaria y la del genético despreciable. Las enfermedades infecciosas producidas por contagio son un ejemplo.
Para la mayoría de los rasgos fenotípicos hay tanto un componente genético como uno ambiental. Muchas veces estamos seguros de la existencia de ambos pero no conocemos, o incluso sabemos que es difícil de estimar, la contribución parcial de cada uno. La mayor parte de las enfermedades no estrictamente hereditarias, como las cardiovasculares, los cánceres, la hipertensión, el asma, el Alzheimer, el Parkinson y las autoinmunes como la artritis y el lupus, pueden tener un componente hereditario, pero sin duda poseen un componente ambiental no despreciable y, a veces, preponderante. En muchos casos la genética molecular ha llegado a descubrir genes cuyas mutaciones provocan predisposición a dichas enfermedades, pero eso no significa que quien tenga esa mutación padecerá indefectiblemente la enfermedad.
El comportamiento humano también es consecuencia de la interacción del genotipo con el ambiente. Sin embargo, salvo en unos pocos trastornos neurológicos hereditarios, no se ha demostrado una influencia genética específica. La pretendida base genética de fenotipos tan complejos como la inteligencia, la orientación sexual, la criminalidad, las capacidades artísticas o deportivas, debe ser tomada con pinzas y sujeta a riguroso análisis experimental en cada caso particular. De lo contrario se corre el riesgo de caer en el determinismo genético, que lejos de ser una ley biológica es un instrumento de discriminación y dominación socioeconómica.
El conocimiento del genoma no autoriza a nadie a estigmatizar a las personas como resultado irreversible de lo que ordenan sus genes. Los genes nos dicen que podemos hablar, pero no qué idioma; que podemos amar, pero no a quién; que podemos disfrutar de la música, pero no de cuál.
Los distintos tipos de inteligencia, las capacidades, los afectos y nuestros actos son resultados del proceso de culturización, el cual no está registrado en ningún gen y, en cambio, está fuertemente influenciado por el ambiente familiar, social y económico en que vivimos.
Lo heredado y lo adquirido:
Para referirnos al carácter heredado o adquirido de un rasgo resulta muy importante diferenciar tres conceptos biológicos: congénito, genético y heredable. Congénito es aquello que le puede ocurrir al embrión o al feto durante la vida intrauterina, sea o no causado por mutaciones en los genes (es decir, genético) y sean o no esas posibles mutaciones heredadas de alguno de los padres (heredable). Así, un defecto o característica congénita de un individuo puede provenir de situaciones vividas por la madre durante el embarazo o simplemente de fenómenos no controlables conocidos como “ruido” del desarrollo embrionario. En este caso, afectará la vida de ese individuo pero no se transmitirá a su descendencia.
Una característica genética está causada por alteraciones en los genes, pero no es necesariamente heredable. Por ejemplo, un tumor de piel tiene origen genético porque está producido por mutaciones en los genes de alguna célula de la piel. Pero ese cambio en los genes de esa célula no es transmitido a la descendencia porque no afecta a las células germinales (óvulos o espermatozoides). Quiere decir que es genético, pero no es heredable.
Por último, lo heredable, que siempre es genético, es lo único que podría ser tenido en cuenta para avalar una teoría puramente determinista. Las alteraciones genéticas se transmiten mediante ciertas leyes de padres a hijos.
Cuando sin pruebas se atribuyen de comportamiento o capacidades intelectuales humanas a los genes, y se supone que las variantes de estos genes están distribuidas en forma asimétrica en distintos grupos humanos, se terminan postulando hipótesis deterministas como, por ejemplo, que ciertos grupos tienen un techo intelectual y que no “vale la pena” invertir dinero en educación para ellos porque están “genéticamente” limitados.
Genoma, variabilidad genómica y razas:
Podemos definir una especie como un conjunto de individuos capaces de dar descendencia fértil por reproducción sexual. Los individuos que pertenecen a una misma especie a menudo presentan diferencias genéticas. Esto es así porque, aunque para cada gen el individuo presenta dos alelos –dos variantes del mismo gen–, el número de alelos de un gen existente en la especie en su conjunto es generalmente mayor que dos. Un gen puede tener decenas de alelos diferentes, pero en cada individuo sólo habrá dos de ellos. Una raza es una subpoblación de individuos de una especie que tiene una alta homogeneidad genética, es decir que los individuos que la componen comparten muchos más alelos entre sí que con cualquier otro individuo de la misma especie, pero de otra raza.
Las comparaciones de secuencias de ADN entre humanos indican que las grandes diferencias genéticas, de existir, tienen lugar entre individuos y no entre poblaciones. En términos más sencillos, por ejemplo, una persona caucásica (blanca) de Europa puede compartir muchas más variantes alélicas con un asiático o un africano que con otro europeo del mismo color de piel. Dos negros africanos pueden distar mucho más genéticamente entre sí que cualquiera de ellos respecto de un blanco. Esto hace, por ejemplo, que en muchos casos la histocompatibilidad (o sea, cuán compatibles son sus órganos y tejidos) entre un negro y un blanco sea mayor que entre dos individuos de la misma “raza” y, como consecuencia, un negro sea más apto que un blanco para donar un órgano de trasplante a otro blanco. Estudios moleculares del genetista Svante Paabo confirman que las razas humanas no existen. El mismo concepto fue expresado en forma sencilla y elegante por el genetista brasileño Sérgio Pena: “No es que seamos todos iguales, sino que somos todos igualmente distintos”.
Una de las fuentes de variabilidad intraespecífica son los cambios de una base en la misma posición entre distintos individuos. Estos cambios son llamados SNPs (pronunciado snips), por single nucleotide polymorphisms. Si bien los SNPs no están uniformemente distribuidos a lo largo de nuestro genoma, se estima que aparece en promedio un SNP cada mil bases de secuencia. Esto indica que el genoma de nuestra especie, Homo sapiens, es altamente homogéneo con una similitud de secuencia del 99,9%. Las similitud de secuencia entre humanos y chimpancés es de alrededor del 98%. No obstante, hay diferencias, tanto entre humanos entre sí como entre humanos y chimpancés que no se deben a SNPs sino a variaciones del número de copias de un dado segmento de ADN dentro del mismo genoma, lo cual puede elevar las diferencias entre individuos a niveles del 5-6%.
Aún no está claro cómo contribuyen las variaciones en el número de copias al fenotipo de los humanos.

--------------------------------------------------------------------------------------------------------------------
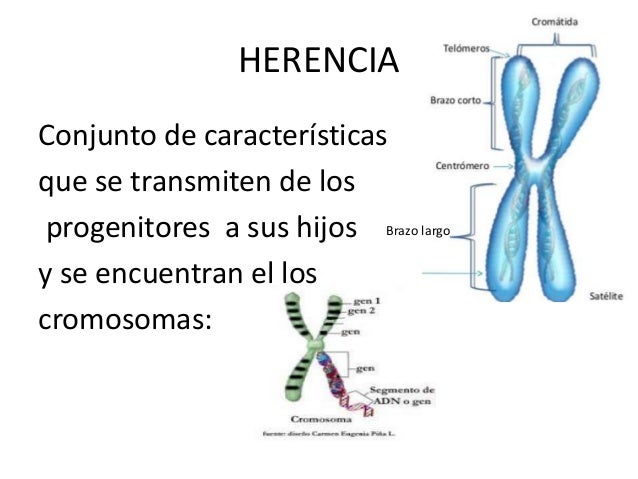
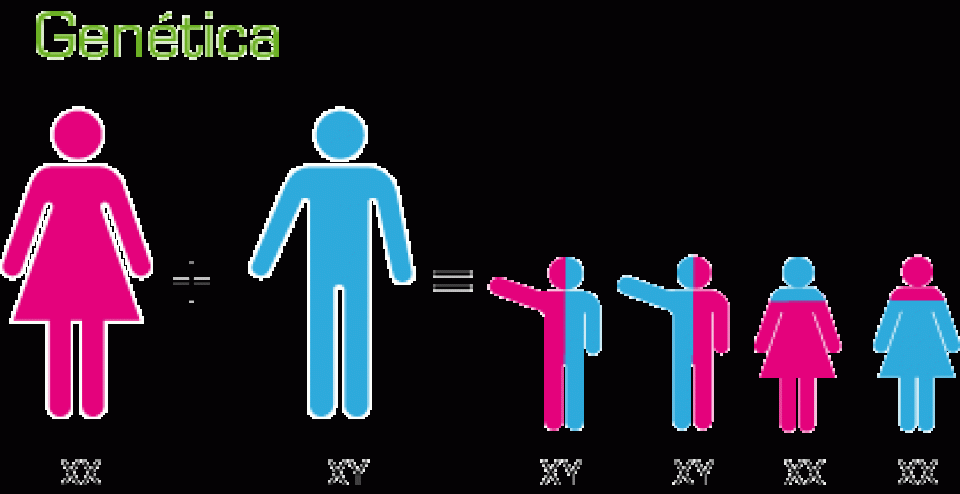
MECANISMOS DE HERENCIA
El primero en realizar experimentos decisivos sobre la herencia y en formular las leyes básicas de la genética fue Gregor Johann Mendel (1822-1884), quien distinguió entre el carácter visible y el factor que determinaba su aparición, el gen. Un gen es la unidad de la herencia que se transmite en un gameto y que rige el desarrollo de un carácter, por interacción con otros genes, el citoplasma y el ambiente.
Naturalmente, puesto que los gametos son los únicos elementos que pasan de los padres a la generación siguiente, en ellos tiene que hallarse el mecanismo de la herencia.
Los genes existen en pares (pares de alelos) en las células de los individuos diplontes, mientras que los gametos sólo tienen un gen de cada tipo.
Un par de alelos está compuesto por el gen paterno y el gen materno de dos cromosomas homólogos. El par de alelos que rige un determinado carácter se simboliza con una pareja de letras iguales (una representa el gen paterno y otra el materno), aunque cada uno de ellos puede hacerlo con una característica diferente (por ejemplo, uno puede regir el color pardo de los ojos y el otro el color azul).
Cuando el par de alelos está formado por dos genes idénticos, y que por tanto se manifiestan de la misma manera (el mismo color de los ojos), se dice que el individuo que los posee es homocigoto para el carácter que determinan; mientras que los híbridos que contienen dos alelos antagónicos son heterocigotos.
La homocigosis se representa con ambas letras, mayúsculas o minúsculas; la heterocigosis, con una mayúscula y otra minúscula. En la heterocigosis, el miembro de un par de alelos que se expresa es el dominante, mientras el otro que sólo se expresaría en condiciones de homocigosis- es el recesivo.
En el caso de herencia dominante las letras mayúsculas representan los genes dominantes y las minúsculas los recesivos. Pero en la heterocigosis también puede haber herencia intermedia o codominancia, cuando dos alelos tengan la misma potencia para determinar el carácter, siendo entonces los heterocigotos de aspecto intermedio al de los progenitores respecto a dicho carácter.
La constitución genética completa de un individuo, tanto la manifiesta como la latente, constituye su genotipo y su aspecto; es decir, el conjunto de caracteres evidentes (morfológicos, fisiológicos, bioquímicos y hasta psicológicos), constituye su fenotipo.
No hay comentarios.:
Publicar un comentario